
- Title
-
Heparan Sulfate Proteoglycans Regulate Fgf Signaling and Cell Polarity during Collective Cell Migration
- Authors
- Venero Galanternik, M., Kramer, K.L., Piotrowski, T.
- Source
- Full text @ Cell Rep.
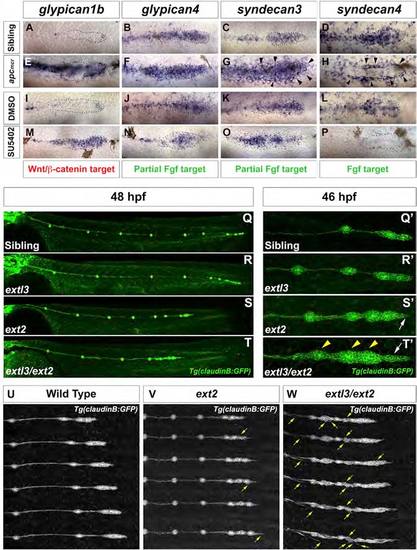
Loss of HSPGs Results in LL Prim Migration and Patterning Defects (A, E, I, and M) gpc1b expression in (A) WT, (E) apcmcr, (I) DMSO-treated, and (M) SU5402-treated embryos. (B, C, F, G, J, K, N, and O) gpc4 and sdc3 expression in (B and C) WT, (F and G) apcmcr, (J and K) DMSO-treated, and (N and O) SU5402-treated embryos. (D, H, L, and P) sdc4 expression in (D) WT, (H) apcmcr, (L) DMSO-treated, and (P) SU5402-treated embryos. (G and H) Black arrowheads point to the ectopic halo of sdc3 and sdc4. (Q and R) WT and extl3 mutant prim has reached the tail tip by 48 hpf. (S and T) ext2 and extl3/ext2 prim fails to complete migration. (Q′-T′) Magnification of prim at 46 hpf. (Q′ and R′) WT and extl3 mutant prim shows a normal morphology. (S′) ext2 mutant prim possesses a pointed tip (white arrow). (T′) extl3/ext2 mutant prim shows severe morphological defects. The leading edge elongates (white arrow) and rosettes are closely spaced (yellow arrowheads). (U-W) Still images of time-lapse movies: WT (U), ext2 (V), and extl3/ext2 mutant prim (W). Yellow arrows in (V) and (W) point to ectopic filopodia. See also Figure S2 and Movies S1, S2, S3, and S4. |

The Wnt/β-Catenin Pathway Is Ectopically Activated in extl3/ext2 Mutants (A-M′) In extl3/ext2 mutants Wnt/β-catenin target genes expand progressively between 34 and 48 hpf. (F, H, J, and L) Sibling prim migration has finished by 48 hpf. (G, I, K, and M) In extl3/ext2 mutants, the prim stalls and lef1, axin2, sef, and fgf10 are upregulated. (G′, I′, K′, and M′) Magnified views of extl3/ext2 mutant prim. Black arrows: Wnt/β-catenin target expression in interneuromast cells. (N-U′) Time course of the Wnt/β-catenin target wnt10a and inhibitor dkk1b. Upregulation of wnt10a correlates with the downregulation of dkk1b. (O, O′, S, and S′) Mild downregulation of dkk1b corresponds with the expansion of wnt10a at 40-42 hpf. (P, P′, T, and T′) By 44-46 hpf, wnt10a expands along the prim and dkk1b expression is almost lost. (Q, Q′, U, and U′) By 48-50 hpf, dkk1b expression is entirely reduced, resulting in complete expansion of wnt10a. (V-Y′′) The Fgf target pea3 is gradually lost in the prim (V-Y′) but is progressively activated in surrounding cells (V′′-Y′′). (Z and Z′) Lower-magnification images of the 44-46 hpf embryos in X, X′, and X′′. See also Figures S2 and S3. |

Fgf Signal Transduction Is Progressively Lost in extl3/ext2 Mutant Prim (A-B′) ZO-1 staining reveals apical constrictions (white arrows) in 48 hpf extl3/ext2 neuromasts and prim rosettes, but not in interneuromast clumps (yellow arrow). (C-D′) LL hair cells precursors are present in 48 hpf extl3/ext2 mutants, as evidenced by atoh1a and s100t expression. Black arrows in (C), (D), and (D′) indicate the position of the prim. (E-H′′) Dp-ERK staining reveals remaining low levels of Fgf signaling in the 48 hpf prim. Dp-ERK is progressively downregulated as rosettes collapse (E′′-H′′). By 5 dpf, no protein remains inside the prim (H′). (I-J′) By 3 dpf, most of the formed hair cells die inside the collapsing neuromast. (I′ and J′) Neuromasts outlined in the white boxes in (I) and (J). (K-L′) Inhibition of Fgf signaling accelerates the onset of the extl3/ext2 phenotype. extl3/ext2 mutants and their siblings treated with SU5402 from 28 to 34 hpf show complete upregulation of the Wnt target lef1. See also Figures S2, S3, and S5 and Movie S4. EXPRESSION / LABELING:
PHENOTYPE:
|

NaClO3 Treatment Phenocopies extl3/ext2 Mutants, Causing Loss of Fgf Signaling and Resulting in Expansion of the Wnt Domain (A and B) In NaClO3-treated embryos, the prim stalls. (C-D′′) NaClO3-treated embryos show downregulation of HS levels by 10E4 antibody staining. (E-F′′) extl3/ext2 mutants show a similar loss of HS. (G-H′) NaClO3-treated Wnt reporter line embryos show expansion of the Wnt domain in the prim, as evidenced by destabilized GFP (dGFP) expression and dgfp in situ hybridization. (I-J′) The Fgf target pea3 is restored in NaClO3-treated prim after CAfgfr1 induction. (K-L′) Restored Fgf signaling restricts expression of the Wnt target lef1 back to the leading region. (M-N′) The restored restriction correlates with the activation of dkk1b after Fgf pathway activation. (O-T′) Wnt is restricted after NaClO3 treatment by inducing Dkk1b expression in the Tg(hsp70:dkk1b-GFP) line. In the absence of HS, Dkk1b induction is sufficient to restrict the Wnt targets lef1 and fgf3 back to the leading region (O-P′ and Q-R′), but not to restore Fgf signaling in the trailing region of the prim (S-T′). pea3 is still only expressed in cells surrounding the prim (T′). See also Figure S4, Table S1, and Movies S5 and S7. |

Chemokine Receptor Expression Correlates with Prim Stalling in extl3/ext2 Mutants (A-H) cxcr4b time course in 32-50 hpf siblings and extl3/ext2 mutants. cxcr4b is upregulated at 32-34 hpf and increases as the embryo develops. (I-P) cxcr7b is expressed normally between 32 and 42 hpf in extl3/ext2 mutants, but becomes downregulated at 44 hpf until it is absent by 50 hpf. These expression changes correlate with the expansion of Wnt signaling ( Figure 2) and slowing down and stalling of the prim. (Q-X) cxcl12a time course shows that despite the upregulation of cxcl12a at 32 hpf, the prim continues to migrate until cxcr7b is downregulated by 44-46 hpf (M-P). cxcl12a upregulation continues as the extl3/ext2 phenotype becomes severe by 50 hpf (W and X). (Y, Y′, and Z) extl3/ext2 mutants possess more Engrailed (4D9)-positive medial fast muscle fibers along the trunk. In (Z) the SE is indicated. See also Figure S4. |

Loss of Fgf Signaling and HS Function Leads to Random, Dynamic Filopodia Formation (A-B′) BIO-treated Tg(claudinB:GFP) embryos show upregulation of cxcl12a compared to control embryos, but no evidence of ectopic filopodia formation. BIO treatment leads to upregulation of the Wnt and Fgf pathways in the prim. (C-D′) In NaClO3-treated Tg(claudinB:GFP) embryos where cxcl12a is upregulated. Filopodia form along all LL cells (D′). Similarly to extl3/ext2 mutants, these embryos show upregulation of Wnt and loss of Fgf signaling. (E-F′) cxcl12a is only slightly upregulated in ext2 mutants. Mild ectopic filopodia are observed in the most anterior region of the prim by time-lapse recording (F′; Movie S3); 48 hpf ext2 mutants show only partial Wnt upregulation and Fgf signaling downregulation (data not shown). (G-H′) Bmp signaling inhibition with Dorsomorphin leads to an increase in cxcl12a-expressing muscle cells. Bmp inhibition does not alter prim migration or signaling, but induces the upregulation of cxcl12a. (I-J′) Inhibition of Fgf signaling by SU5402 causes the formation of ectopic filopodia, but not upregulation of cxcl12a (J′). Wnt is upregulated as a result of Fgf inhibition. (K and L) Inhibition of Fgf signaling in extl3/ext2 mutants does not eliminate ectopic protrusions. Black arrows point to ectopic filopodia. See also Figures S4 and S5 and Movies S6 and S8. |

Wnt/β-catenin and Fgf signaling regulate LL primordium migration and patterning. Summary of previously published data. (A) Diagram depicting the position of the LL migrating primordium at 24 hpf in a WT embryo, followed by a magnified view of the primordium showing the Wnt/β-catenin and Fgf signaling domains with their respective target genes. Modified from (Aman and Piotrowski, 2008). |

extl3/ext2 mutants show a late onset phenotype. Related to Figure 1-3. (A-B′′) extl3/ext2 mutants show significant downregulation of HS chains as evidenced by 3G10 antibody whole mount staining. (C-F) LL phenotypes in 5 dpf extl3/ext2 mutants. Compared to ext2 and extl3/ext2 mutants, extl3 mutants show only mild defects in the LL at this stage of development (D). ext2 mutant primordia do not reach the tail tip by 5 dpf and possess an elongated tip but clear neuromast formation (E). extl3/ext2 mutants are characterized by a collapsed LL where all neuromasts have lost their round shape, likely due to the loss of Fgf signaling (F). (G-H′) Wnt/β-catenin targets lef1 and fgf3 are expanded along the whole lateral line in 3 dpf extl3/ext2 mutants. (I-K′) The Fgf target fgfr1 is progressively downregulated between 34 hpf and 3 dpf in extl3/ext2 mutants. (K′) Close up of a 48 hpf extl3/ext2 primordium showing the downregulated fgfr1 expression. |

Pharmacological inhibition of HS recapitulates extl3/ext2 mutants. Related to Figures 2-4. (A-C′) The ectopic pea3 expression surrounding the primordium is Fgf dependent in extl3/ext2 mutants (B-B′) and NaClO3-treated embryos (C-C′). (D-F′′) The ectopic pea3 expression is restricted to cells immediately surrounding the primordium in extl3/ext2 mutants (E-E′′) and NaClO3-treated embryos (F-F′′) as demonstrated by double labeling with ClaudinB:GFP and pea3. (G-H′′) 3G10 labels HS chains and its persistence in NaClO3 treated embryos shows that only HS chain modifications are lost (See Table S1). (I-W′) Time course of the Fgf targets pea3 and dkk1b and the Wnt/β-catenin target lef1 in DMSO controls compared to NaClO3-treated embryos. Similar to extl3/ext2 mutants, lef1 expands only as pea3 and dkk1b are gradually downregulated after NaClO3- treatment. |

Hh signaling is upregulated in extl3/ext2 mutants and cxcl12a overexpression does not induce ectopic filopodia. Related to Figure 5-6. (A-D) In situ hybridization of sibling and extl3/ext2 mutant embryos at 36 hpf shows the expression of the Shh target gene patched1 along the trunk (A and B) and magnified views of the notochord in the same fish in A′ and B′. patched1 stays upregulated along the tail trunk of extl3/ext2 mutants at 44 hpf (C and D), especially around the notochord (C′ and D′) where Shh signaling is active. (E-F) Tg(claudinB:GFP) fish were crossed to Tg(hsp70:sdf1a) fish and heat shocked to overexpress cxcl12a. Images were taken after heat shock (~34 hpf). A rounded primordium was observed in transgenic siblings with mild ectopic filopodia formation at the leading edge. (G-J) The Wnt/β-catenin target lef1 is expressed in the leading region of the primordium at 34 hpf (G) but a short 3 hourtreatment with the GSK inhibitor BIO (2 µM) leads to its expansion along the entire primordium (H). The chemokine receptor cxcr7b is expressed in the trailing region of the primordium at 34 hpf (I) and it is inhibited by Wnt/β-catenin signaling after BIO treatment resulting in primordium stalling (J). hpt: Hours post treatment. |
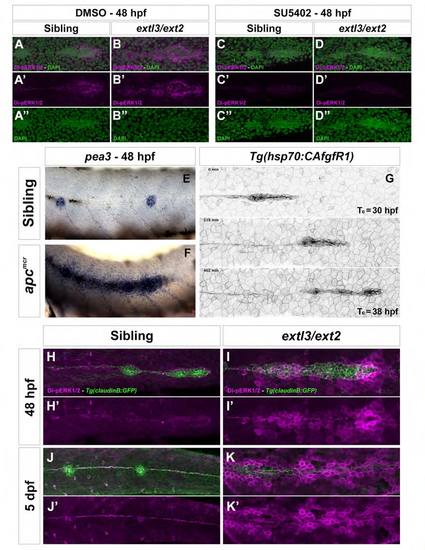
Wnt/β-catenin upregulation in apcmcr mutants also leads to an Fgfdependent ectopic halo of pea3. Related to Figure 3 and Figure 6 (A-D′′) A 4-hour treatment of SU5402 inhibits Di-pERK1/2 in cells surrounding the primordium in sibling (A, C) and mutant embryos (B, D), demonstrating that ERK is Fgf dependent. (E-F) pea3 expression in siblings (E) and apcmcr mutant (F) embryos at 48 hpf. In apcmcr mutants the stalled primordium activates pea3 expression ectopically in cells surrounding the primordium. (G) Heat shock induced constitutive activation of Fgf signaling does not lead to ectopic filopodia formation in the lateral line. (H-K′) extl3/ext2 mutant embryos hyper-activate Di-pERK1/2 in primordium surrounding tissues like the skin compared to siblings. This phenotype is evident at 48 hpf (D-E′) and becomes more pronounced by 5 dpf (F-G′). |