
- Title
-
Rescue of bmp15 deficiency in zebrafish by mutation of inha reveals mechanisms of BMP15 regulation of folliculogenesis
- Authors
- Zhai, Y., Zhang, X., Zhao, C., Geng, R., Wu, K., Yuan, M., Ai, N., Ge, W.
- Source
- Full text @ PLoS Genet.
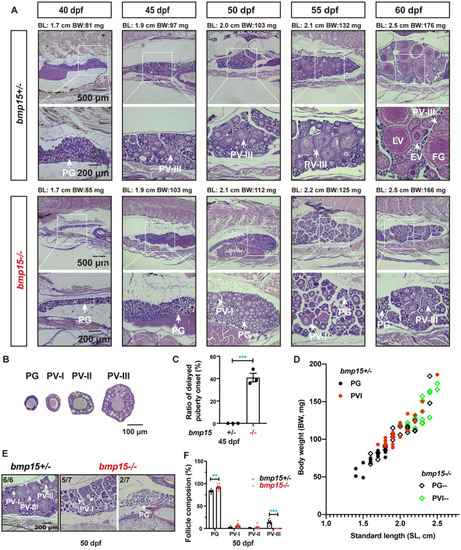
Delayed follicle activation and puberty onset in (A) Phenotype analysis of the early follicle development and PG–PV transition in control ( |
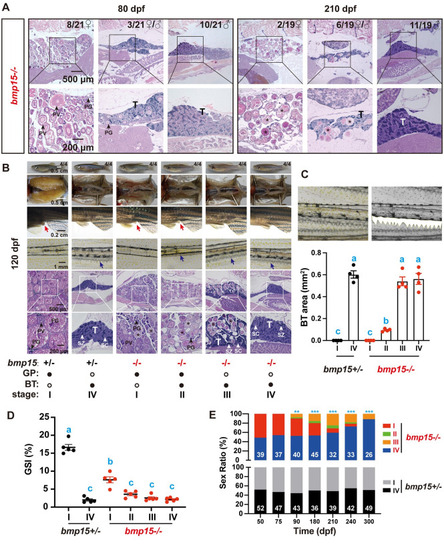
Sex reverse in (A) Histology analysis of the mutant ( |

Partial rescue of (A) Histological examination of gonads at 120 dpf. The follicle development was arrested at PV–EV transition in |

Transcriptome analysis of (A) Heatmap of DGEs among four genotypes of |

Expression of selected genes at PG and PV stages in (A) Stage-matching ovarian samples collected at 60 dpf. Both PG and PV follicles were present in |
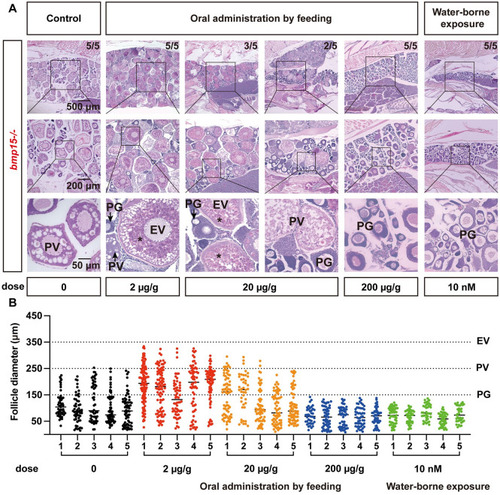
Rescue of vitellogenic growth in (A) Histology analysis of the |

Effect of fadrozole on vitellogenic growth in (A) Morphology and histology of the double mutant gonads ( |

Role of (A) Morphology and histology of the gonads in different genotypes at 120 dpf. The asterisk shows vitellogenic follicles without yolk mass in the triple mutant ( |

Expression of selected genes in different genotypes at 120 dpf. (A) Expression of ovarian genes in the PG follicles ( |

Interaction of (A) Histological analysis of |
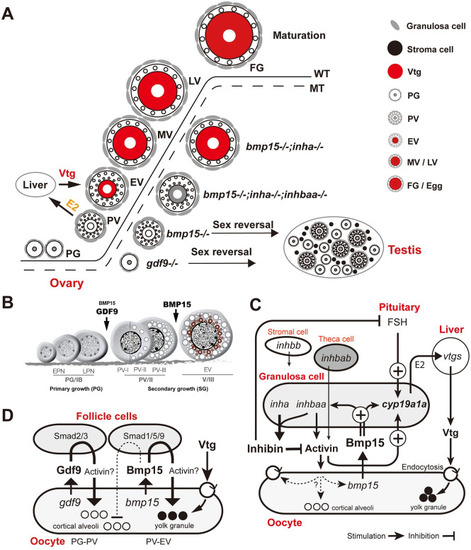
Summary of genetic analysis and working hypotheses on mechanisms of Bmp15 action in zebrafish. (A) Zebrafish folliculogenesis and phenotypic defects of single ( |