
- Title
-
The entire zebrafish blastula-gastrula margin acts as an organizer dependent on the ratio of Nodal to BMP activity
- Authors
- Fauny, J.D., Thisse, B., and Thisse, C.
- Source
- Full text @ Development
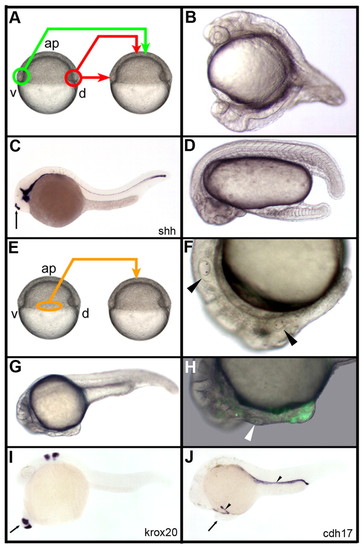
Organizing properties of the early gastrula margin. (A) Dorsal (red) and ventral (green) marginal grafts of the early gastrula embryo. (B) Transplantation from the dorsal to ventral margin organizes secondary head and trunk. (C) A graft of the dorsal margin at the animal pole induces a short stretch of notochord labeled with the sonic hedgehog shh probe (arrow). (D) A graft of the ventral margin at the animal pole results in the formation of a secondary tail. (E-H) Grafts of the lateral margin (orange, E) organize partial secondary axes containing anterior trunk and posterior head structures (arrowheads in F, otic vesicles in endogenous and secondary axes) that extend from the endogenous head (G) and contain both GFP-labeled cells from the donor, as well as unlabeled cells from the host (H; arrowhead, secondary otic vesicle). (I,J) In situ hybridizations using krox20 and cdh17 RNA probes. Arrow in I,J, secondary axes; arrowhead in J, pronephric ducts; ap, animal pole. B-D,F-J, lateral view, anterior to the left, dorsal to the top; A,E, lateral view, anterior to the top. |
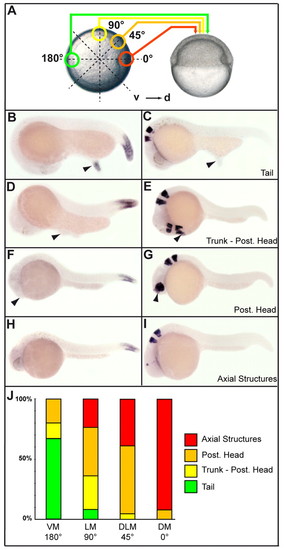
Continuous organizing properties of the early gastrula margin. (A) Various gastrula marginal grafts performed at the animal pole of host embryos. Donor embryo, left, animal pole view; host embryo, right, lateral view. 0°, dorsal margin (DM); 45°, dorsolateral margin (DLM); 90°, lateral margin (LM); 180°, ventral margin (VM). (B-I) In situ hybridizations for hoxC12b (B,D,F,H) and for krox20 (C,E,G,I) on embryos displaying secondary tails (B,C), a trunk-posterior head (D,E), a posterior head (F,G) and axial structures (H,I). Arrowheads indicate secondary axes. (J) The frequency of the different structures induced by grafts of the various pieces of margin. Number (n) of embryos analyzed: DM, 15; DLM, 18; LM, 24; VM, 15. B-I, lateral view, anterior to the left, dorsal to the top. |
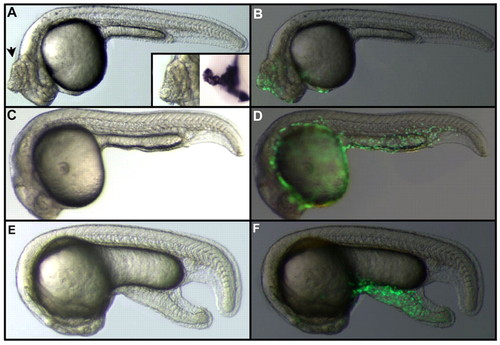
Effect of Nodal or BMP on animal pole blastomeres. (A) Injection of nodal RNA in an animal pole blastomere at the 128- to 256-cell stage results in the formation of ectopic axial structures (arrow) that express shh, a marker of the notochord and floor plate (inset). (B) Localization of the clone derived from the injected blastomere visualized by fluorescence resulting from the translation of gfp RNA co-injected with nodal RNA. (C) Injection of bmp4 RNA in an animal pole blastomere at the 128- to 256-cell stage has no effect on the morphology of the embryo. (D) Cells derived from the injected blastomere are all located in the epidermis. (E) Injection of nodal and bmp4 RNAs in an animal pole blastomere at the 128- to 256-cell stage results in the formation of an ectopic tail. (F) Localization of the clone derived from the injected blastomere visualized by fluorescence resulting from the translation of gfp RNAs co-injected with nodal and bmp4 RNAs. |
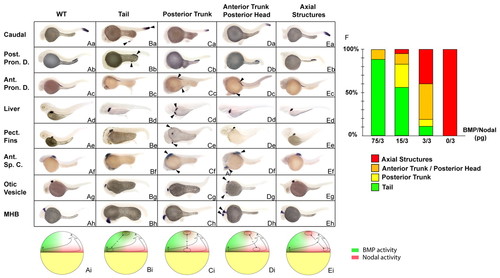
Simultaneous stimulation of Nodal and BMP activities recapitulates the organizing activities of the different marginal domains. (Aa-Eh) In situ hybridization with markers of caudal structures (hoxC13b, first row), posterior pronephric ducts (Post. Pron. D., slc12a3, second row), anterior pronephric ducts (Ant. Pron. D., slc4a2, third row), liver (fabp10, fourth row), pectoral fins (Pect. Fins, si:dkey-70p6.3p, fifth row), anterior spinal cord (Ant. Sp. C., rxrγ, sixth row), otic vesicles (starmaker, seventh row) and the midbrain-hindbrain boundary (MHB, engrailed 2, eighth row) on embryos co-injected with nodal and a range of bmp RNAs in an animal pole blastomere at the 128-cell stage and sorted based on the morphology of the induced secondary structures, as indicated at the top. WT embryo (Aa-h), embryo displaying a secondary tail (Ba-h), posterior trunk (Ca-h), anterior trunk/posterior head (Da-h) and axial structures (Ea-h). (Ai-Ei) Schematic representation of embryos at late blastula stage, dorsal to the right, depicting the type of marginal grafts at the animal pole of a host blastula that result in the formation of the structures indicated above the figure. Green, the ventral domain corresponding to high levels of BMP; red, the activity of Nodal at the blastula margin. (F) The frequency of the different classes generated after injection of various dilutions of bmp and nodal RNAs. Green, tail; orange, posterior trunk; yellow, anterior trunk/posterior head; red, notochord; WT, wild type; arrowheads, endogenous and ectopic structures formed. Lateral view. For each condition, at least 120 embryos were injected in two independent experiments. EXPRESSION / LABELING:
|
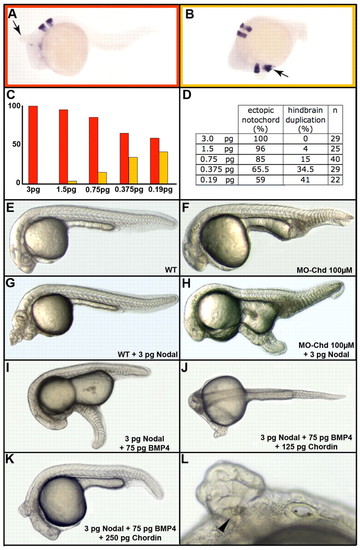
Interaction between endogenous BMP activity and ectopic Nodal signaling at the animal pole. (A-L) Injection of 3 pg nodal RNA at the animal pole results in the induction of ectopic axial structures (A, arrow). When lower amounts are injected, partial secondary axes extend posteriorly up to the hindbrain (B, arrow). The bar graph in C, built from data presented in D, presents the percentage of notochord (red) or hindbrain (orange) duplication as a function of the amount of nodal RNA injected. Owing to the presence of endogenous BMP activity at the animal pole, the progressive decrease of nodal RNA mimics the BMP/Nodal ratio present from the dorsal to the lateral margin. n, number of embryos analyzed. Wild type (WT, E) and chordin morphant (MO-Chd, F) embryos. Injection of 3 pg nodal RNA at the animal pole of WT embryos results in the formation of an ectopic notochord (G), whereas injection of the same amount in the chordin morphant results in formation of a secondary tail (H). Injection of 3 pg nodal and 75 pg bmp4 RNAs into wild type embryos results in formation of a secondary tail (I). Co-injection of 125 pg chordin RNA changes this posterior structure into a secondary trunk (J), whereas co-injection of 250 pg chordin RNA results in the formation of posterior head structures (K,L, with a secondary otic vesicle, arrowhead). Lateral view, anterior to the left and dorsal to the top. |
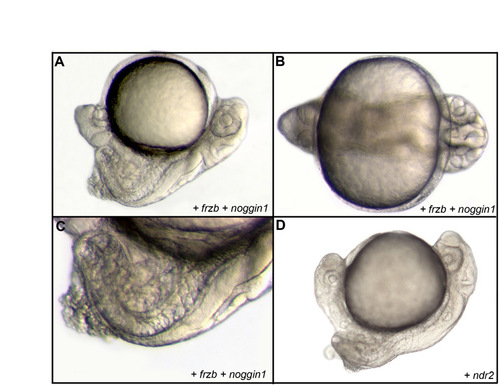
Formation of complete secondary embryonic axes. (A-D) Formation of a complete secondary axis, extending anteriorly up to the anterior head and containing axial tissue, after injection of noggin1 and frzb RNAs into a ventral blastomere at the 16-cell stage in lateral (A) and frontal (B) views. (C) Detail of the trunk region in lateral view. An identical secondary axis can be obtained upon overexpression of nodal by injection of ndr2 RNA into a ventral blastomere at the 16-cell stage, as shown in lateral view (D). |
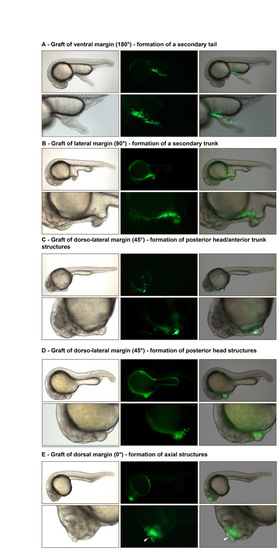
Organizer activity of the lateral blastula-early gastrula margin. (A-E) Grafts of various parts of the early gastrula margin from a donor labeled with GFP to the animal pole of a host blastula (unlabeled) result in the organization of a partial ectopic secondary axis that contains labeled (fluorescent, green) cells from the donor as well as unlabeled cells that have been recruited from the animal pole of the host. (A) Graft of the ventral margin. (B) Graft of the lateral margin. (C,D) Grafts of the dorso-lateral margin resulting in the formation of posterior head/anterior trunk structures (C) or posterior head (D). (E) Graft of the dorsal margin resulting in the formation of a secondary axial structure (arrowhead, unlabeled notochord cell). Lateral view, anterior to the left, dorsal to the top. Left panels, live embryo; middle panels, fluorescently labeled cells; right panels, merge of live and florescent images. |