
- Title
-
A point mutation decouples the lipid transfer activities of microsomal triglyceride transfer protein
- Authors
- Wilson, M.H., Rajan, S., Danoff, A., White, R.J., Hensley, M.R., Quinlivan, V.H., Recacha, R., Thierer, J.H., Tan, F.J., Busch-Nentwich, E.M., Ruddock, L., Hussain, M.M., Farber, S.A.
- Source
- Full text @ PLoS Genet.
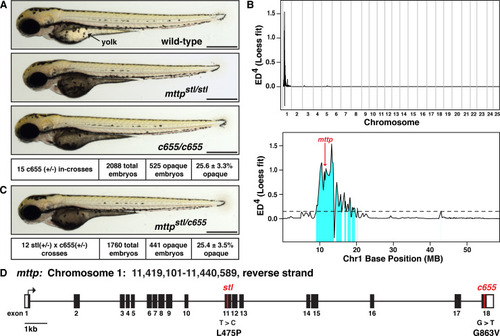
(A) Representative images of a wild-type zebrafish embryo, a homozygous mutant embryo carrying the previously described PHENOTYPE:
|

(A) (Top) Cartoon depicting the cross-sectional view of a 4 dpf zebrafish embryo. The YSL surrounds the yolk mass and serves as the embryonic digestive organ. The dashed box indicates the view expanded in the bottom panel and in panel B. (Bottom) Stored yolk lipids undergo lipolysis in yolk platelets (YP), presumably releasing free fatty acids into the YSL. These fatty acids are re-esterified in the ER bilayer to form TG, PL, and cholesterol esters. The lipids are packaged into B-lps in the ER with the help of Mtp and are likely further processed in the Golgi before being secreted into the perivitelline space (PS) and then circulation. (B) Representative transmission electron micrographs of the yolk and YSL from wild-type and PHENOTYPE:
|

(A) LipoGlo fish express the NanoLuc luciferase enzyme as a C-terminal fusion on ApoBb.1 as a result of TALEN-based genomic engineering [ |
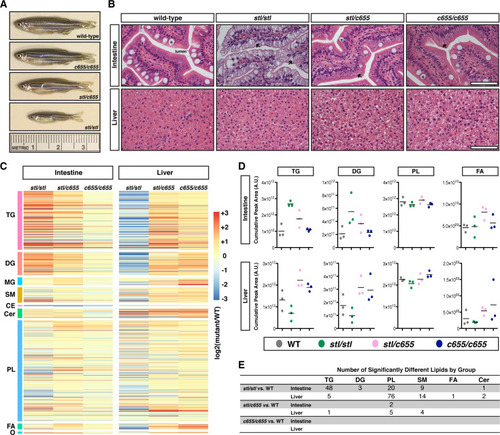
(A) Representative images of male WT and PHENOTYPE:
|

(A, B) COS-7 cells were first transfected with an expression vector for human APOB48 (5 μg), distributed equally in 6-well plates, and subsequently transfected with plasmids expressing either wild-type zebrafish |
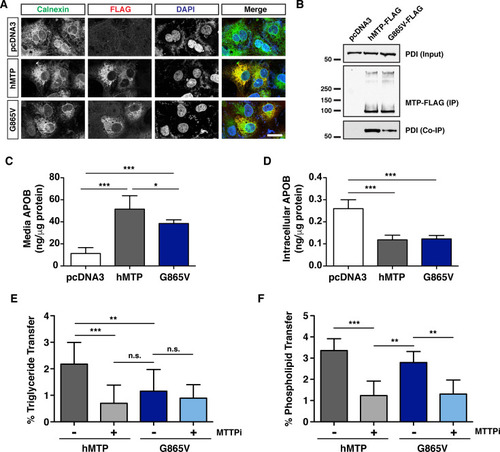
(A) Immunofluorescence in COS-7 cells expressing wild-type human MTTP-FLAG or human MTTP(G865V)-FLAG proteins using anti-FLAG (red) and anti-Calnexin (green) antibodies; scale = 25 μm. (B) Human MTP-FLAG proteins (WT and G865V) were immunoprecipitated from COS-7 cell lysate (400 μg protein) using the M2 flag antibody and immunoblots were probed for both FLAG and PDI. For input, 15 μg protein was used. (C, D) COS-7 cells were co-transfected with human APOB48 and either wild-type human |
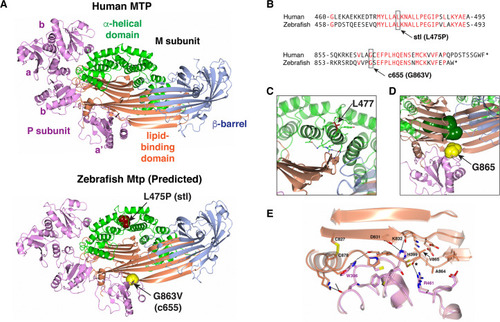
(A) Ribbon representation of the human MTP complex (PDB entry 6I7S) and the Zebrafish modeled structure. The positions of L475 and G863 in the Zebrafish structure are shown in space-filling representation. (B) Alignment of human MTP and zebrafish Mtp amino acid sequences surrounding the |