
|
Fig 8
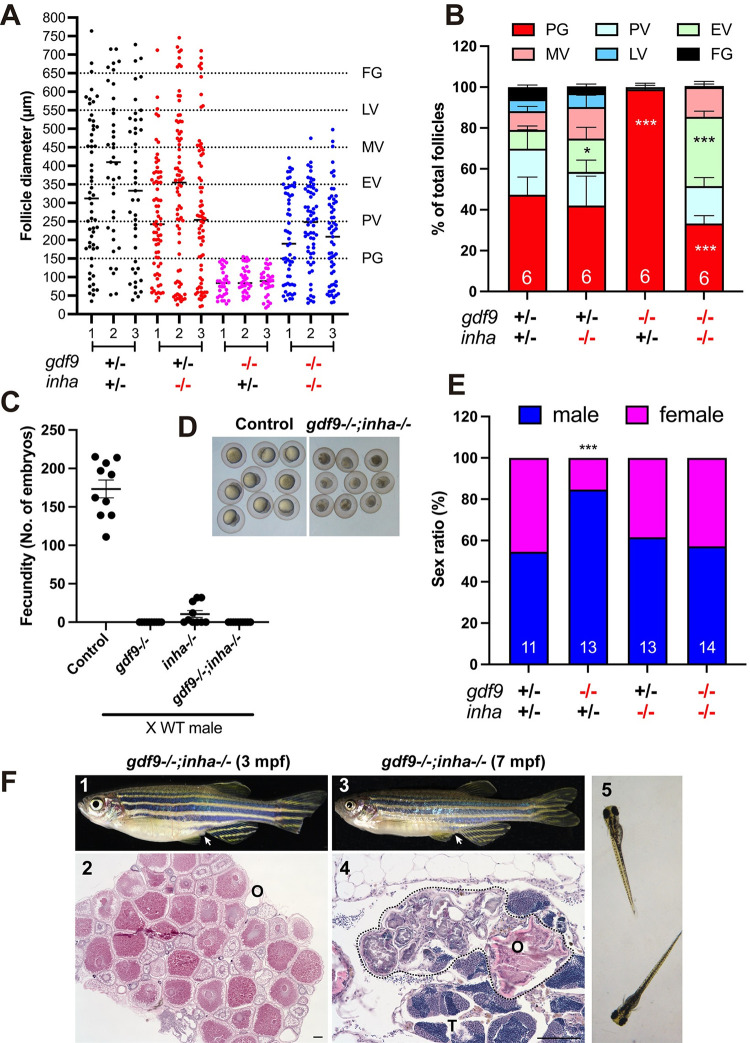
(A and B) Follicle composition in different genotypes at 90 dpf. The follicles in gdf9-/- were arrested at PG stage. Although the PG-PV transition and yolk accumulation resumed in the double mutant (gdf9-/-;inha-/-), the vitellogenic growth could not proceed to the FG stage, and the follicles were arrested at the MV-LV transition. * P < 0.05; *** P < 0.001 (X2 test, n = 6). (C) Fertility test on females of single (gdf9-/- and inha-/-) and double mutants (gdf9-/-;inha-/-) at 90 dpf. Five mutant and control females were tested 10 times consecutively every other day by natural pair breeding with WT males (n = 5/test). The test was conducted over about 20 days. In each test, the number of fertilized eggs or embryos was counted for each female and the data point represents the average of the five fish tested. (D) Eggs spawned by the control (gdf9+/-;inha+/-) and double mutant (gdf9-/-;inha-/-). The eggs from the double mutant were smaller than those from the control. (E) Prevention of sex reversal in gdf9-/- by double mutation with inha-/- at 90 dpf. The gdf9-/- fish showed a male-biased sex ratio (82% males), and this was corrected in the double mutant (47% males). *** P < 0.001 (X2 test, n = 11–14). (F) Sex reversal of the gdf9 and inha double mutant (gdf9-/-;inha-/-) at 7 mpf. (1) Double mutant female at 3 mpf with female-specific genital papilla (arrow); (2) Ovary of 3-mpf double mutant; (3) Sex reversed intersexual fish at 7 mpf with regressed genital papilla (arrow); (4) Ovotestis of 7-mpf double mutant. The follicles in the ovarian tissue (O) were degenerating (circled by dotted line) and the testicular tissue (T) became dominant in the gonads. (5) The offspring from sex reversed double mutant males.